
Ümarussid

| |
| Taksonoomia | |
| Riik |
Loomad Animalia |
| Hõimkond |
Ümarussid Nematoda |
Ümarussid ehk nematoodid (Nematoda) on loomade hõimkond. Kirjeldatud on üle 20 000 liigi. Hinnangute järgi on liike kokku 100 000 kuni 10 miljonit; liigirikkuse poolest on ümarussid lülijalgsete kõrval üks liigirikkamaid selgrootute hõimkondi[1] Ümarussid on tõenäoliselt kõige isendirikkam loomarühm: ühe hinnangu järgi kuulub sellesse 80% loomade isenditest[2].
Ümarussid on enamasti suhteliselt väikesed valged kuni värvitud niitjad ussid, kes elavad niiskes keskkonnas. Enamik liike toitub mikroobidest, kuid on ka kiskjaid ning arvukalt (teadaolevalt 15 000) parasiteerivat liiki, kelle peremehed on taimed ja loomad (sealhulgas inimene).
Ümarussid on kohastunud peaaegu kõikide mage- ja merevee- ning maismaaökosüsteemidega, sealhulgas maakoore ülaosa sügavamate piirkondadega ja polaarpiirkondadega. Neid on leitud isegi Antarktikast ja ookeanisüvikutest. Paljudes elupaikades on nad nii isendite kui ka liikide arvu poolest suurim loomahõimkond.
Eesti nematoodidest on ülevaate teinud Eino Krall.
Nematoodide seast varbuss (Caenorhabditis elegans) on bioloogide üheks tähtsaks mudelorganismiks – teda on uuritud erakordselt põhjalikult.
Anatoomia[muuda | muuda lähteteksti]

1 Suuava
2 Sool
3 Kloaak
4 Erituselund
5 Munand
6 Närvisüsteemi neeluvõru
7 Selgmine peanärvitüvi
8 Kõhtmine peanärvitüvi
9 Erituspoor
Kehaehitus[muuda | muuda lähteteksti]
Ümarussid on sind triploblastsed esmassuused. Neil on tüüpiliselt usjas kuju, nad on pikad ja ümmarguse ristlõikega. Lülilisus puudub. Kehaõõs on kitsas pseudotsööl, nagu ka mitmel teisel selgrootute hõimkonnal, mille esindajad on valdavalt usjad ja väga väikesed.
Ümarussi eesotsas on suur lihaseline suuava, mis läheb üle neeluks. Suu ja neelu anatoomia on mitmeti kohastunud toitumisviisiga. Voolikukujulises kehas on seede- ja suguelundid. Pärak asetseb tagaotsast pisut eespool.
Suurim ümaruss on kašeloti platsentas elav Placentonema gigantissima. Emased kasvavad kuni 8,40 m pikkuseks läbimõõduga kuni 2,5 cm, isased kasvavad kuni 4 m pikkuseks läbimõõduga kuni 0,9 cm. See liik kuulub klassi fasmiidsed (Secernentea) (alamklass Spiruria, sugukond Tetrameridae).
Epidermis ja kutiikula[muuda | muuda lähteteksti]
Ümarussi epidermis (naharakukiht) ei koosne üksikutest rakkudest nagu teistel loomadel, vaid rakumaterjali massist, mis ei ole rakumembraanidega rakkudeks jaotatud ja millel on mitu rakutuuma. Sääraseid moodustisi nimetatakse süntsüütsiumiks.
Epidermis eritab tunduvalt paksema, mitmekihilise eksoskeleti – kutiikula, mis kaitseb ümarusse kuivamise või teiste ebasoodsate keskkonnatingimuste eest, parasiitsete liikide puhul ka peremeesorganismi seedemahlade eest. Kuumaveeallikates elavate liikide kõrval on leitud ka liike, mille kutiikula võimaldab taluda pH väärtusi 2,5-st (näiteks sidrunimahl, äädikas) kuni 11,5-ni (näiteks majapidamisammoniaak) või jääda mitu tundi ellu vedelas heeliumis (umbes −272 °C kuni −268 °C). See, et neil on "jäik" kutiikula ja pikilihastik (ringlihased ümarussidel peaaegu puuduvad), võimaldab neil kulgeda ainult siueldes.
Kutiikula koosneb ümarussidel kuni neljast kihist:
- Sisemine kiukiht koosneb diagonaalselt vastassuundades kulgevatest kiududest. Sellest kihist sõltub kutiikula kindlus ja elastsus kõige rohkem.
- Maatriksikihil on vähem määratletud ehitus.
- Kortikaalkiht koosneb kollageenist.
- Väline epikutiikula sisaldab lipiide ja mõnel perekonnal katab seda veel lipiidikiht.
Lihased[muuda | muuda lähteteksti]
Ümarussidel kui ümarloomadel on kulgemiseks ainult pikilihased, mis ulatuvad peast sabani. Lihasrakud koosnevad kolmest osast:
- monotsüüdist, mis on mittekokkutõmbuv osa, mis sisaldab rakutuumi, mitokondreid ja Golgi aparaati;
- kokkutõmbuvast osast, mis sisaldab aktiini- ja müosiinikiude;
- jätkest, lihasrakkude mittekokkutõmbuvast osast, mis saab teiste lihasrakkude või närvidega ühendustesse astuda.

Ümarussi lihased asetsevad torutaoliselt naha all. Seda erinevatest kudedest koosnevat seina nimetatakse nahklihasmõiguks. Tugev välistoes kutiikula ja pseudotsöölivedeliku kõrge siserõhk (70 kuni 210 mmHg) on hüdroskelett. Koos pikilihaste kui antagonistidega saab ümaruss siueldes kulgeda või osa kehaga kõrgele sirutuda.
On ka ringlihased, kuid ainult suu ja päraku juures.
Närvisüsteem[muuda | muuda lähteteksti]
Rõngusside närvisüsteem on väga lihtsa ehitusega. See koosneb neeluvõrust või söögitoruvõrust, millest tahapoole kulgevad selgmine ja kõhtmine peatüvi. Närvisüsteem on võimeline aistima ja töötlema mitmesuguseid lihtsaid ärritajaid. Pikinärvid on lihasrakkude ja tsütoplasmaga tihedas kokkupuutes ja kulgevad läbi kogu keha. Erinevalt teistest loomadest, kellel neuronid levivad lihasteni, levivad lihasrakud ise närviteedeni.
Suguelundid[muuda | muuda lähteteksti]
Suguelundid koosnevad emastel vulvast keha keskel, kusjuures keha lõpposa nihkumine ettepoole on ümarusside apomorfia.
Isastel on keha lõpus moodustunud kloaak, milles on seemnejuha, pärasool ja spikulaaraparaat. Viimane on ümarusside paaritumiselund, mis koosneb paarilistest kõvastunud konksukujulistest spiikulistest, mis paiknevad kutiikula taskus ja mille liikumist juhib gubernaculum. Harva on taandarengu või liitumise tõttu spiikuleid ainult üks. Spiikulite ülesanne ei ole mitte seemne transport, vaid need haagivad isase paaritumise ajal emase vulvasse, nii et spermatosoidid saavad üle minna otse seemnejuhast.[3][4] Enamikul sugukonna Protostrongylidae esindajatel koosneb paaritumiselund kutiikula paksendist kõhtmises ja külgmises kloaagiseinas, mida nimetatakse telamoniks[5]
Füsioloogia[muuda | muuda lähteteksti]
Toitumine[muuda | muuda lähteteksti]
Toit on erinev ning varieerub vabalt elavatel liikidel bakteritest ja vetikatest ning seentest, raipest ja fekaalidest saakloomadeni. Suu juures on sageli väikesed jätked, mida kasutatakse söömiseks või kompimiseks. Seal tõmmatakse toit sisse ja lömastatakse tugevate lihastega. Toit satub sealt siis lihtsasse pikka soolde, kus seda töödeldakse ja seeditakse. Ümarussidel pole ringesüsteemi, millega toitaineid kehas jaotada. Selleasemel töödeldakse toitaineid sooles ja need juhitakse sealt otse läbi seinte keharakkudeni, kus neid tarbitakse.
Sooles võib olla ka endosümbiootilisi mikroorganisme (baktereid ja seeni), mida on tarvis teatud toitainete lõhustamiseks, näiteks tselluloosi katabolismiks. Mõnedelt vähestelt liikidelt, näiteks männi-laguuss ja mardikates elav Pristionchus pacificus, on leitud endogeenseid tsellulaase. Tsellulaasigeenid on saadud horisontaalse geeniülekande teel endosümbiontidelt[6][7].
Hingamine[muuda | muuda lähteteksti]
Hapiniku vastuvõtmine toimub sarnaselt seedimisega. Et ümarussidel pole hingamiselundeid ega ringesüsteemi, võetakse dihapnikku vasti läbi naha ja difundeeritakse otse keharakkudeni.
Paljunemine[muuda | muuda lähteteksti]
Paljunemine toimub suguliselt, enamasti kahe lahutatud sooga. Isased on tavaliselt emastest väiksemad ja neil on sageli iseloomulik kõver sana. On siiski ka iseviljastuvaid hermafodiite, näiteks varbuss. [8] Parasiitsetel liikidel on sageli keeruline elu- ja paljunemistsükkel metageneesiga, millega võib kaasneda peremeesorganismi vahetus või elundi vahetus peremeesorganismis. Seejuures võivad pistesääsklased mikrofilaariad, st ussi vastsed vastu võtta ning anda need järgmisel arenguastmel edasi lõppperemeestele[9].
Kestumine[muuda | muuda lähteteksti]
Ümarussid kestuvad. Sellepärast ja RNA uuringute põhjal arvatakse nad esmassuuste seas kestloomade (Ecdysozoa) ülemhõimkonda. Vabalt elavad ümarussid arenevad enamasti otseselt ning kestuvad kasvu jooksul neli korda.
Elupaigad[muuda | muuda lähteteksti]
Ümarussid esinevad peaaegu kõikjal, meres, magevees ja maismaaökosüsteemides. Neid peetakse üldiselt "sagedasteks ja omnipresentseteks"[10] ja sageli on nad ökosüsteemis esindatud suurema arvu liikide ja isenditega kui teised loomarühmad. Väga olulised on nad mullas, kus nad katavad mitu alumist troofilist taset.
Ümarussid on alustanud ka ekstreemseid elupaiku. Liik Halicephalobus mephisto leiti Lõuna-Aafrikast lõhevetest kuni 3,6 km sügavusest ning on teadaolevalt kõige sügavamal maakoores elav loom[11] Teised ümarussid elavad Antarktises McMurdo kuivorgude muldades, mis on aeg-ajalt väga külmad, soolased ja kuivad, nii et mullavesi pole kättesaadav; nad elavad säärased perioodid üle anhüdrobioosis[12][13]
On ka palju parasiitseid liike, nii taimedes (näiteks peedi-kiduuss) kui ka loomades, sealhulgas inimestes. Inimese sooles elavate parasiitsete ümarusside seas on solge, piuglane Trichuris trichiura mediina niituss, naaskelsaba ja ürgpihtlane. Seevastu filaariad ehk süstikussid Wuchereria bancrofti, Brugia malayi ja Loa loa elavad lümfisoontes või nahaaluses rasvkoes.
Enamik vabalt elavaid ümarusse on mikroskoopilised ja kuuluvad meiofaunasse. Ainult mõned loomaparasiidid kasvavad palju suuremaks.
Parasiitsete liikide nakatamisteed ja -strateegiad[muuda | muuda lähteteksti]

Lõppperemeheks olevad imetajad, sealhulgas inimene, võivad nakatuda, süües toorest liha, milles juba on vastsed (näiteks keeritsussid), või süües ussimune sisse koos roojaga (näiteks koerte puhul). Võidakse ka süüa ussimune sisaldava roojaga saastunud toiduaineid (kui toidutaime väetatakse virtsaga või kui pärast roojamist ei pesta käsi). Paljude liikide puhul on võimalik ka vastsete sissetung läbi naha (näiteks Ancylostoma duodenale, Necator americanus). Mõned filaariad, näiteks jõepimeduse ehk onkotsertsiaasi tekitaja Onchocerca volvulus, levivad putukapistete kaudu.
Mõnd liiki parasiitsed ümarussid suudavad oma peremeesorganismide käitumist või välimust muuta. Eriti keerukas näide on Myrmeconema neotropicum.[14] See uss teeb nii, et tema vaheperemeheks oleva sipelga Cephalotus atratus kõht (gaster), mis on täis selle ussi mune, muutub mustast punaseks. Nakatatud sipelgad, kes erinevalt nakatamata liigikaaslastest ei ilmuta ohu puhul agressiivset kaitsekäitumist ega tooda alarmaineid, sirutavad oma punast tagakeha üles. Tõenäoliselt peab see ligi meelitama viljadest toituvaid linde, kes on ussi lõppperemehed, ning söövad ära sipelgate kõhud, mida on ülejäänud kehast kerge lahti rebida[15].
Männi-laguuss kasutab forentidena puidusikkusid[16][17].
Taksonoomia[muuda | muuda lähteteksti]
Hõimkonna autor on Nathan Cobb (1919), kes pani sellele nimeks Nemata (vanakreeka sõnast νῆμα 'niit').
Hiljem vaadeldi ümarusse (Nematoda) seltsina nüüd enam mitte kehtivas ümarloomade hõimkonnas.
Praegu peetakse ümarusse jälle hõimkonnaks (Nematoda või Nemata).
Süstemaatika[muuda | muuda lähteteksti]
- Fasmiiditud (Adenophorea)
- Fasmiidsed (Secernentea)
eElurikkus annab niisuguse klassifikatsiooni:
- perekond Choronema
- perekond Iotalaimus
- perekond Macfadyenia
- perekond Notosouthernia
- perekond Triplonchium
- sugukond Marimermithidae
- sugukond Neotonchidae
- sugukond Rhadinematida
- klass Adenophorea
- sugukond Chrongastridae
- klass Chromadorea
- selts Araeolaimida
- sugukond Axonolaimidae
- sugukond Bodonematidae
- sugukond Camacolaimidae
- sugukond Coninckiidae
- sugukond Creagrocercidae
- sugukond Cylindrolaimidae
- sugukond Diplopeltidae
- sugukond Halaphanolaimidae
- sugukond Haliplectidae
- sugukond Ohridiidae
- sugukond Pakiridae
- selts Ascaridida
- sugukond Atractidae
- sugukond Cucullanidae
- sugukond Kathlaniidae
- sugukond Quimperiidae
- sugukond Raphidascarididae
- sugukond Seuratidae
- sugukond Toxocaridae
- selts Chromadorida – silmlaik-ümarussilised
- perekond Rhabdocoma
- sugukond Achromadoridae
- sugukond Benthimermithidae
- sugukond Ceramonematidae
- sugukond Choanolaimidae
- sugukond Chromadoridae
- sugukond Cyatholaimidae
- sugukond Diplopeltoididae
- sugukond Ethmolaimidae
- sugukond Hypodontolaimidae
- sugukond Selachinematidae
- sugukond Tarvaiidae
- selts Desmodorida
- sugukond Choniolaimidae
- sugukond Desmodoridae
- sugukond Draconematidae
- sugukond Epsilonematidae
- sugukond Microlaimidae
- sugukond Monoposthiidae
- sugukond Richtersiidae
- selts Desmoscolecida
- sugukond Cyartonematidae
- sugukond Desmoscolecidae
- sugukond Myliidae
- selts Diplogasterida
- selts Monhysterida
- perekond Cyartonema
- sugukond Aponchiidae
- sugukond Comesomatidae
- sugukond Fusivermidae
- sugukond Linhomoeidae
- sugukond Monhysteridae
- sugukond Scaptrellidae
- sugukond Siphonolaimidae
- sugukond Sphaerolaimidae
- sugukond Xyalidae
- selts Oxyurida
- sugukond Hystrignathidae
- sugukond Pharyngodonidae
- sugukond Pseudonymidae
- sugukond Thelastomatidae
- sugukond Travassosinematidae
- selts Plectida – ürglehtersuulised
- sugukond Aulolaimidae
- sugukond Bastianiidae
- sugukond Euteratocephalidae
- sugukond Leptolaimidae
- sugukond Odontolaimidae
- sugukond Plectidae
- sugukond Rhabdolaimidae
- sugukond Teratocephalidae
- selts Rhabditida – varbussilised
- perekond Agamonema
- perekond Amphicaecum
- perekond Paracanthocheilus
- perekond Torynurus
- sugukond Myolaimidae
- sugukond Ancylostomatidae
- perekond Ancylostoma – kõõrpea
- sugukond Bunonematidae
- sugukond Cylindrocorporidae
- sugukond Diplogasteridae
- sugukond Diplogasteroididae
- sugukond Diploscapteridae
- sugukond Heterorhabditidae
- sugukond Metastrongylidae
- sugukond Neodiplogasteridae
- sugukond Odontopharyngidae
- sugukond Pseudodiplogasteroididae
- sugukond Pterygorhabditidae
- sugukond Rhabdiasidae
- sugukond Rhabditidae
- sugukond Rhabditidae
- sugukond Rhabditinematidae
- sugukond Spiruridae
- sugukond Strongylidae
- sugukond Trichostrongylidae
- sugukond Tylopharyngidae
- sugukond Acuariidae
- sugukond Anisakidae
- sugukond Ascarididae – solgelased
- perekond Angusticaecum – angustitsääk
- perekond Ascaris
- liik Ascaris lumbricoides – inimesesolge
- liik Ascaris suum – seasolge
- perekond Toxocara
- liik Toxocara canis – kutsikasolge
- liik Toxocara cati – kassisolge
- sugukond Ascaridiidae
- sugukond Camallanidae
- selts Araeolaimida
Artikli kirjutamine on selles kohas pooleli jäänud. Jätkamine on kõigile lahkesti lubatud. |
Kivistised[muuda | muuda lähteteksti]

Ümarussidel ei ole kehaosi, mis säiliksid kaua ja oleksid settekivimites hästi äratuntavad. Sellepärast on nende kivistisi vähe ja ainult teatud laadi.[18][19]
Enamik ümarusside kivistisi ja kõige paremini säilinud kivistised on leitud kriidi ja tertsiaari merevaigust, kusjuures vanimad eksemplarid pärinevad Liibanoni Alam-Kriidist[18][20][21]. Vanimad otsesed kivistised pärinevad Šotimaalt Alam-Devoni Rhynie Cherti leiukohast ja on ühtlasi vanimad otsesed tõendid parasiitsetest ümarussidest. Need koosnevad munadest, juveniilsetest staadiumidest ja täiskasvanud isenditest, mis avastati 2005 varajase maismaataime Aglaophyton major õhulõhealusest õõnest ja mida kirjeldati 2008 nime Palaeonema phyticum all.[18] Vabalt meres elavate ümarusside jälgfossiilidena tõlgendatud settestruktuure kirjeldati Hiina Hubei provintsi Alam-Ordoviitsiumi savikividest[19].
Siberist Kolõma madalikust leiti ümarusse, kes olid umbes 46 000 aastat tagasi igikeltsas konserveerunud. Hoolimata sellest, et nad olid hilispleistotseenist saadik külmunud, olevat kaks isendit, kes identifitseeriti kui Panagrolaimus aff. detritophagus ja Plectus aff. parvus, edukalt elustatud[22][23].
Pilte[muuda | muuda lähteteksti]
-
 Solge (Ascaris lumbricoides)
Solge (Ascaris lumbricoides) -

-

-
 Heterodera glycines ja muna
Heterodera glycines ja muna -
 Keeritsussid (Trichinella spiralis)
Keeritsussid (Trichinella spiralis) -
 Mononchidae söömas oma liigikaaslast
Mononchidae söömas oma liigikaaslast -

-
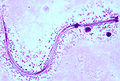


_penetrates_a_tomato_root_-_USDA-ARS.jpg)





Vaata ka[muuda | muuda lähteteksti]
Viited[muuda | muuda lähteteksti]
- ↑ George O. Poinar, Jr. Phylum Nemata. – James H. Thorp, D. Christopher Rogers (toim). Keys to Nearctic Fauna. Thorp and Covich's Freshwater Invertebrates, Volume II, 4. trükk, Academic Press/Elsevier, 2016, ISBN 978-0-12-385028-7, lk 169–180, siin lk 169. Viide allikale: G. O. Poinar, Jr. The Evolutionary History of Nematodes. Brill: Leiden 2001.
- ↑ T. Bongers, H. Ferris. Nematode community structure as a bioindicator in environmental monitoring. – Trends Ecol Evol, kd 14, nr 6, 1999, lk 224–228. Resümee.
- ↑ Estelle V. Balian jt. Freshwater Animal Diversity Assessment, Springer 2008, ISBN 978-1-4020-8258-0, lk 68–69.
- ↑ Peter Ax. Multicellular Animals: Order in Nature – System Made by Man: 3, Springer 2003, ISBN=978-3-540-00146-1, lk 19–20.
- ↑ Wörterbuch der Veterinärmedizin, 2. trükk, lk 1196.
- ↑ John T. Jones, Cleber Furlanetto, Taisei Kikuchi. Horizontal gene transfer from bacteria and fungi as a driving force in the evolution of plant parasitism in nematodes. – Nematology, 2005, kd 7, nr 5, lk 641–646. Resümee.
- ↑ Werner E. Mayer, Lisa N. Schuster, Gabi Bartelmes, Christoph Dieterich, Ralf J. Sommer. Horizontal gene transfer of microbial cellulases into nematode genomes is associated with functional assimilation and gene turnover. – BMC Evolutionary Biology, 2011, kd 11, nr 1, lk 13. Veebis.
- ↑ Liigil Steinernema longicaudum tekib isasel alles siis küps suguaparaat, kui emane on läheduses, vt: L. Ebssa, I. Dix, C. Griffin. – Current Biology, kd 18, nr 21, lk R997–R998
- ↑ Filarien, Lexikon der Biologie, Spektrum Akademischer Verlag 1999, vaadatud 13.9.2023
- ↑ Gregor W. Yeates, Howard Ferris, Tom Moens, Wim H. Van der Putten. The Role of Nematodes in Ecosystems. – Michale J. Wilson, Thomas Kakouli-Duarte (toim). Nematodes as environmental indicators, CAB International 2009, ISBN 978-1-84593-385-2, lk 1–44.
- ↑ G. Borgonie, A. García-Moyano, D. Litthauer, W. Bert, A. Bester, E. van Heerden, C. Möller, M. Erasmus, T. C. Onstott. Nematoda from the terrestrial deep subsurface of South Africa. – Nature, 2011, kd 474, nr 7349, lk 79–82. [1], doi:10.1038/nature09974. Vt ka: Dave Mosher. New "Devil Worm" Is Deepest-Living Animal. – National Geographic News, 2. juuni 2011.
- ↑ Amy M. Treonis, Diana H. Wall, Ross A. Virginia. The use of anhydrobiosis by soil nematodes in the Antarctic Dry Valleys. – Functional Ecology, 2000, kd 14, nr 4, lk 460–467. Veebis.
- ↑ Amy M. Treonis, Diana H. Wall. Soil nematodes and desiccation survival in the extreme arid environment of the Antarctic Dry Valleys. – Integrative and Comparative Biology, 2005, kd 45, nr 5, lk 741–750. Veebis.
- ↑ George O. Poinar, Jr., Stephen P. Yanoviak. Myrmeconema neotropicum n. g., n. sp., a new tetradonematid nematode parasitising South American populations of Cephalotes atratus (Hymenoptera: Formicidae), with the discovery of an apparent parasite-induced host morph. – Systematic Parasitology, 2008, kd 69, nr 2, lk 145–153. Resümee.
- ↑ Stephen P. Yanoviak, Michael E. Kaspari, Robert Dudley, George O. Poinar, Jr. Parasite-Induced Fruit Mimicry in a Tropical Canopy Ant. – American Naturalist, 2008, kd 171, nr 4, lk 536–544. Resümee.
- ↑ Ute Schönfeld. Der Kiefernholznematode (Bursaphelenchus xylophilus). Pflanzenschutzinformation: Pflanzengesundheitskontrolle 54/2015. Pflanzenschutzdienst des Landes Brandenburg, Landesamt für Ländliche Entwicklung, Landwirtschaft und Flurneuordnung, Frankfurt (Oder) 2015. Veebis.
- ↑ Therese Plüss, Simone Prospero, Thomas Röthlisberger, Bea Schwarzwälder, Christiane Lellig (toim). Leitfaden zum Umgang mit dem Kiefernholznematoden (Bursaphelenchus xylophilus), Eidgenössischer Pflanzenschutzdienst, Bundesamt für Umwelt, Bundesamt für Landwirtschaft, Bern 2015. Veebis.
- ↑ 18,0 18,1 18,2 George O. Poinar, Jr. The Geological Record of Parasitic Nematode Evolution. – Kenneth De Baets, D. Timothy J. Littlewood (toim). Fossil Parasites. Advances in Parasitology, kd 90, Elsevier 2015, lk 53-92. Resümee.
- ↑ 19,0 19,1 Andrzej Baliński, Yuanlin Sun, Jerzy Dzik. Traces of marine nematodes from 470 million years old Early Ordovician rocks in China. – Nematology, 2013, kd 15, nr 5, lk 567–574. Resümee.
- ↑ George O. Poinar, jr., Aftim Acra, Fadi Acra. Earliest fossil nematode (Mermithidae) in cretaceous Lebanese amber. – Fundamental and Applied Nematology, 1994, kd 17, nr 5, lk 475–477.
- ↑ Ülevaade Läänemere merevaigu leidudest eotseenis: Wolfgang Weitschat, Wilfried Wichard. Atlas der Pflanzen und Tiere im Baltischen Bernstein, Pfeil 1998, ISBN 978-3-931516-45-1, lk 54–56.
- ↑ A. V. Shatilovich, A. V. Tchesunov, T. V. Neretina, I. P. Grabarnik, S. V. Gubin, T. A. Vishnivetskaya, T. C. Onstott, E. M. Rivkina. Viable Nematodes from Late Pleistocene Permafrost of the Kolyma River Lowland. – Doklady Biological Sciences, 2018, kd 480, nr 1, lk 100–102. Resümee.
- ↑ Fadenwürmer überleben 46.000 Jahre im Permafrost, mdr.de.
Välislingid[muuda | muuda lähteteksti]
| Pildid, videod ja helifailid Commonsis: Ümarussid |